Spatial modeling of sexual and non-sexual traits
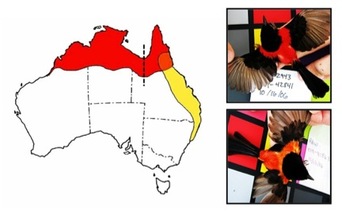
During speciation, diverging populations will likely begin to differ in multiple types of morphological traits. Morphometric traits that describe body shape and size should vary according to the physical environment if these traits are important to survival. In contrast, sexual signals might diverge stochastically with little or no environmental influence. I used the spatial modeling technique generalized dissimilarity modeling (GDM) to quantify variation in sexual and non-sexual morphological between two subspecies of the Red-backed Fairy-wren. This technique compares the relative influence of bioclimatic environmental variables (e.g., temperature, rainfall, vegetation) and geographic distance on phenotypic variation.
GDM analyses revealed that the two subspecies exhibited variation in numerous non-sexual traits (e.g., weight, wing length, tarsus length) that was well explained by underlying environmental variation, but was not broadly diagnostic of the two subspecies. One putative sexual signal, tail length, exhibited a similar pattern, but with a small influence of geographic distance, suggesting it has diverged stochastically to some extent, but is also constrained by the environment. In stark contrast, variation in plumage color (quantified by reflectance spectrophotometry) was not well explained by environment, but rather exhibited a clear pattern of isolation by distance, suggesting it has diverged stochastically as a result of divergent Fisherian sexual selection while the subspecies were geographically isolated. In addition, there was no appreciable divergence in any morphological traits across the genetic barrier in northwest Queensland (the dashed vertical line on the map), while divergence in plumage color was most pronounced across the plumage contact zone in northeast Queensland (the intermediately-colored area on the map). This pattern suggests that alleles for red plumage may have introgressed eastward across the hybrid zone following secondary contact.
GDM analyses revealed that the two subspecies exhibited variation in numerous non-sexual traits (e.g., weight, wing length, tarsus length) that was well explained by underlying environmental variation, but was not broadly diagnostic of the two subspecies. One putative sexual signal, tail length, exhibited a similar pattern, but with a small influence of geographic distance, suggesting it has diverged stochastically to some extent, but is also constrained by the environment. In stark contrast, variation in plumage color (quantified by reflectance spectrophotometry) was not well explained by environment, but rather exhibited a clear pattern of isolation by distance, suggesting it has diverged stochastically as a result of divergent Fisherian sexual selection while the subspecies were geographically isolated. In addition, there was no appreciable divergence in any morphological traits across the genetic barrier in northwest Queensland (the dashed vertical line on the map), while divergence in plumage color was most pronounced across the plumage contact zone in northeast Queensland (the intermediately-colored area on the map). This pattern suggests that alleles for red plumage may have introgressed eastward across the hybrid zone following secondary contact.
